8 The Merger
Imagine you are a chemist, and you have exactly one test tube.
Not one test tube on the bench while your equipment cupboard holds dozens more. One test tube – that’s it. Every reaction you run has to happen in that single vessel. Acid and base, oxidation and reduction, synthesis and degradation – all dumped together in the same pot, all at the same time. Whatever you make is immediately exposed to whatever else is in there. If one reaction produces something fragile, the next reaction may destroy it before you can use it.
That is the prokaryotic cell. For roughly two billion years, bacteria and archaea ran the planet’s chemistry inside a single compartment: the cytoplasm. To be fair, some had a second compartment – gram-negative bacteria carry a periplasmic space between their two membranes. So call it two test tubes for those. “But two tubes is certainly not enough for a good chemical laboratory!”1
And yet prokaryotes thrived. They invented photosynthesis, nitrogen fixation, sulfur cycling, methanogenesis. They reshaped the atmosphere and the ocean floor. They did extraordinary chemistry with minimal architecture. (Some prokaryotes did evolve limited internal membranes – cyanobacteria house their photosynthetic machinery in thylakoid folds, and a few other lineages have specialized compartments – but these are exceptions. Most prokaryotic chemistry still runs in a single well-mixed volume.)
But there were things they could not do. They could not run an oxygen-consuming reaction in one room while running an oxygen-sensitive reaction next door. They could not isolate their DNA behind a membrane and control who got access. They could not build a body made of trillions of differentiated cells, because differentiation requires compartments within compartments – a bureaucracy of nested enclosures, each with its own chemistry, its own imports and exports, its own protected interior.
For that, you need more test tubes. Many more.
The question is: how do you get them?
8.1 The compartment problem
The answer that evolution found—at least for the two compartments that mattered most—is so strange it took biologists a century to accept it. You do not evolve your power plant or your solar panel from scratch. One cell takes up residence inside another – and stays.
This is the story of the eukaryotic cell – the kind of cell that makes up every animal, plant, fungus, and protist on Earth. It is the story of the most consequential merger in the history of life. And it began not with a mutual agreement but with one cell ending up inside another.
The word “eukaryote” means “true kernel” – a reference to the nucleus, the membrane-bound compartment where the genome lives. But the nucleus is only the most visible upgrade. Look inside a eukaryotic cell under an electron microscope and you find a landscape of internal membranes: the endoplasmic reticulum folded into sheets and tubes, the Golgi apparatus stacking its cisternae, lysosomes loaded with digestive enzymes, peroxisomes handling dangerous oxidation reactions, and – most crucially for our story – mitochondria and, in photosynthetic lineages, chloroplasts.
Most of this internal architecture – the nucleus, the ER, the Golgi, the lysosomes – appears to have arisen from the host cell’s own membranes, elaborated through invagination and specialization. But the two organelles that changed the game – mitochondria and chloroplasts – arrived from outside: captured cells that never left.
Each of these compartments is a separate reaction chamber. Each maintains its own internal chemistry, buffered from the rest of the cell by a lipid bilayer. The endoplasmic reticulum can fold proteins under conditions that would wreck the cytoplasm’s redox balance. Lysosomes can run acid hydrolysis at pH 5 while the cytoplasm holds steady at pH 7. Mitochondria can maintain a proton gradient across their inner membrane precisely because that membrane is sealed – a private reservoir of electrochemical potential, insulated from the larger cell.
Compartmentalization buys you the ability to run incompatible processes simultaneously, each sealed behind its own membrane. A laboratory with as many test tubes as you need.
The prokaryotic cell had none of this internal architecture. It was the open-plan office of biology – everything in one room, everyone hearing everyone else’s conversations. Efficient, in a way. Fast, certainly. But limited in the complexity of chemistry it could orchestrate.
8.2 A bridge between worlds
For a long time, the origin of eukaryotes was a black box. Prokaryotes on one side, eukaryotes on the other, and a vast gulf of cellular complexity between them. Then, in 2015, a team of researchers pulled something remarkable out of the Arctic Ocean.
The samples came from deep-sea sediments near a hydrothermal vent field called Loki’s Castle, on the Mid-Atlantic Ridge between Norway and Greenland, at a depth of 3,283 meters.2 The organisms they found were not eukaryotes. They were archaea – prokaryotes, single-celled, lacking nuclei and internal membranes. But their gene set told a different story.
These archaea, dubbed Lokiarchaeota, carried genes that no one expected to find in a prokaryote.3 Genes for actin-like cytoskeletal proteins – the molecular scaffolding that eukaryotic cells use to change shape, crawl, and engulf particles. Genes suggesting the capacity for membrane remodeling. Genes whose eukaryotic counterparts participate in phagocytosis – the act of wrapping your cell membrane around another object and pulling it inside. Whether these archaea actually engulfed particles remains debated.
Lokiarchaeota did not have a nucleus. They did not have mitochondria. But they had the genetic toolkit that could, in principle, lead to both. They were closer to eukaryotes than any other prokaryote ever found – a bridge between the two great domains of cellular life.
Lokiarchaeota was the first discovered member of what is now called the Asgard superphylum – a group of archaea named after figures from Norse mythology (Loki, Thor, Odin, Heimdall).4 Phylogenetic analyses consistently place eukaryotes within the Asgard archaea, not as their sister group.5 This means that eukaryotes did not diverge from archaea; they emerged from within them. The “three domains of life” model (Bacteria, Archaea, Eukarya) may need to be revised to a “two domains” model, with eukaryotes as a highly derived branch of the Archaea. The discovery did not answer every question about eukaryotic origins, but it narrowed the search space dramatically.
8.3 A rival story about the nucleus
The mitochondrion is firm. The nucleus is not. Few events in cell evolution carry evidence as clean as the alpha-proteobacterial origin of mitochondria. The nucleus draws rival explanations.
One of them gives viruses a central role. Takemura argues that giant-virus factories offer a plausible template for the nucleus because they separate DNA replication from ribosome-rich cytoplasm and control traffic across that boundary.6 Medusavirus sharpened the case by carrying histones and other signs of long exchange with eukaryotic lineages. The proposal remains a minority view. It belongs in the chapter because it widens the cast: viral infection may have shaped not only gene traffic in microbial worlds but the architecture of the eukaryotic cell itself.
8.4 Genes that travel without bodies
Long before one cell moved inside another for good, prokaryotes had already evolved a looser form of merger. They exchange genes sideways.
Biologists call this horizontal gene transfer: DNA moving by transformation, conjugation, transduction, and the quieter mechanisms that make microbial evolution look less like a clean tree and more like a web.7 In bacteria, the effect is enormous. A lineage can acquire an enzyme set, a resistance cassette, or an entire metabolic shortcut without waiting for those genes to evolve from scratch. Viral infection spreads genes. Plasmids spread genes. Pieces of dead cells spread genes. The microbial world is full of partial mergers that stop at the genome.
Even minerals get involved. DNA adsorbed to clay and oxide surfaces can survive longer than naked DNA in water, turning sediments into reservoirs of genetic possibilities.8 That ties evolution back to geomicrobiology. Rock does not only host microbial evolution. Minerals can help move metabolic innovation through the biosphere.
By the time endosymbiosis appears, biology is already comfortable with hybridization. Whole-cell merger is unusual. Gene traffic is not.
Somewhere around 1.6 to 2.7 billion years ago – with most molecular clock estimates favoring roughly 2 billion years ago – an archaeal cell, perhaps something like Lokiarchaeota or a close relative, took a step that prokaryotes had been building toward for billions of years.910 It may have been able to reshape its membrane using actin-like scaffolding. How the association with the bacterium began – whether by engulfment, by the archaeon extending membrane protrusions around it, or by the bacterium invading – remains one of the open questions in evolutionary biology.11 However it started, the bacterium was not digested.
That bacterium was an alpha-proteobacterium – an aerobic organism that could use oxygen to burn organic molecules with extraordinary efficiency. Inside the archaeal host, the bacterium kept breathing. It kept producing ATP. And over time, what started as a captured meal became something else entirely: a permanent resident, a co-dependent partner, and eventually an organelle.
That organelle is the mitochondrion. Every mitochondrion in every eukaryotic cell on Earth descends from that single, ancient partnership.
8.5 The spectrum of integration
The transformation from free-living bacterium to mitochondrion did not happen overnight. It was a long, slow slide from partnership to dependence to irreversible fusion – a process that took hundreds of millions of years and that we can still watch happening today, frozen at different stages in different organisms.
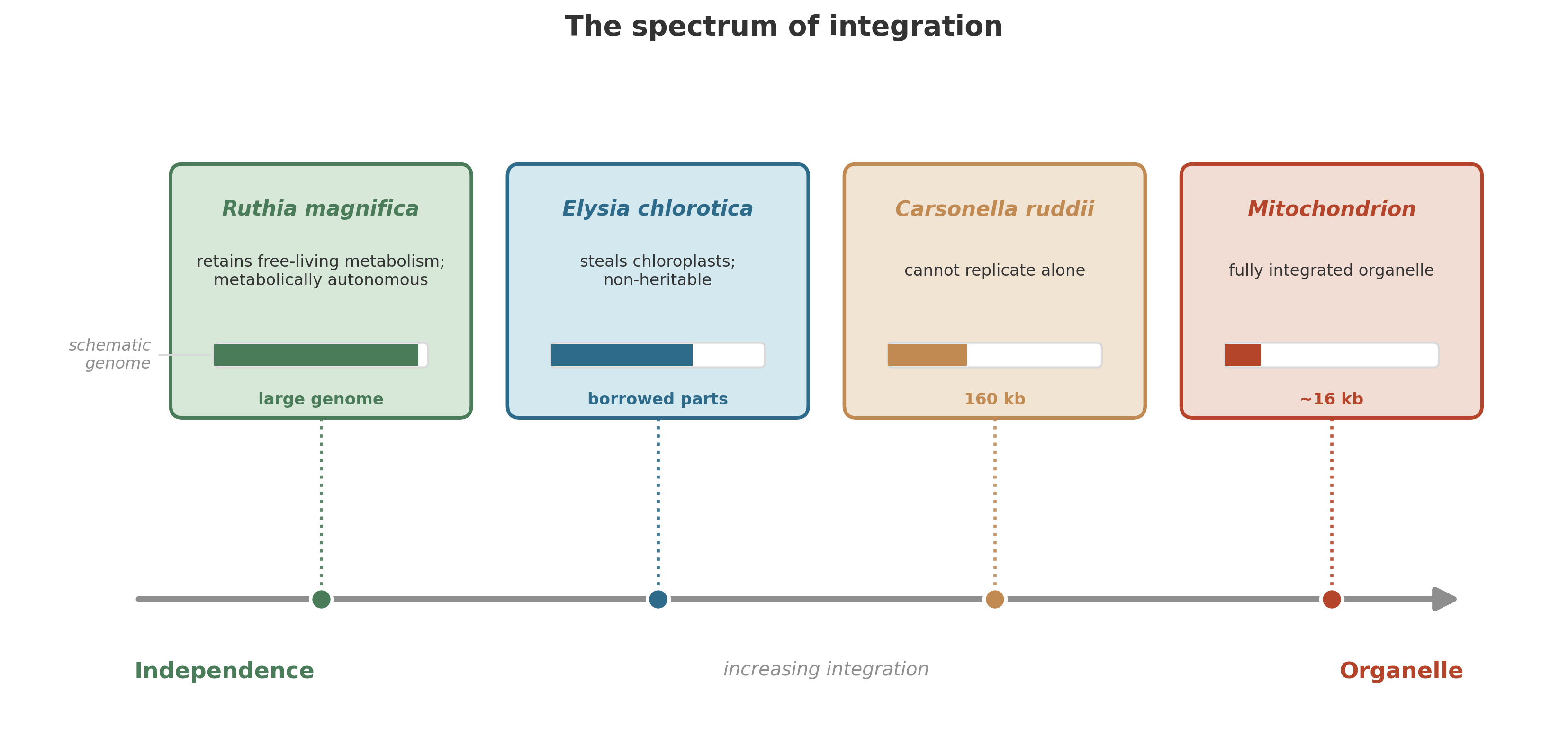
Think of it as a spectrum. At one end, a bacterium lives inside a host cell but retains its full genetic and metabolic independence: it could, in principle, be extracted and grown on its own. At the other end, the bacterium has lost so many genes that it is no longer an organism at all – it is an organelle, a part of the host, unable to exist independently. Between these extremes lies every shade of partnership, dependence, and dissolution.
The living world is full of symbioses caught at different points on this spectrum. They illustrate, at different stages, how symbiotic dependence can deepen – though only a few have crossed (or will cross) the threshold to true organelle status.
8.5.1 The chemist in the dark: Ruthia magnifica
The ocean floor, several kilometers below the surface, is usually a desert. No light penetrates. There is no photosynthesis. The only food drifting down from the productive surface waters is a thin, unreliable drizzle of organic particles – “marine snow” – barely enough to sustain the sparse communities of the abyssal plain.
But where the Earth’s crust is cracked, where hydrothermal fluids seep upward carrying hydrogen sulfide and methane, the desert blooms. Around these vents and seeps, life is dense, improbable, and vivid: tube worms with blood-red plumes, ghostly white shrimp, and clusters of giant clams pressing their tissues against the chemical-rich water.
The giant clam Calyptogena magnifica is one of these vent animals. It lives along the edges of hydrothermal vents on the ocean floor, and its secret is inside its gill cells.12 There, packed into specialized host cells called bacteriocytes, lives Ruthia magnifica – a gamma-proteobacterium that has traded the open ocean for a captive existence inside an animal.
Ruthia is a chemoautotroph. It fixes carbon from CO\(_2\) via the Calvin cycle, just as a plant does, but it does not use sunlight as its energy source. Instead, it oxidizes sulfur compounds. Hydrogen sulfide flows in from the vent fluid; Ruthia strips electrons from it, storing elemental sulfur in intracellular granules, then oxidizing the sulfur further to sulfite and sulfate, extracting energy at each step. That energy drives carbon fixation, and the organic molecules Ruthia produces feed the clam.
The clam, in turn, provides Ruthia with a stable physical environment, a supply of oxygen (delivered via the clam’s blood), and access to the hydrogen sulfide welling up from below. It is a partnership: the bacterium is the chemist, the clam is the house.
What makes Ruthia remarkable is what its genome reveals: it has retained a complete set of genes for chemoautotrophic life.13 It can still, in principle, do everything a free-living sulfur-oxidizing bacterium can do. Its genome has not yet suffered the erosion that afflicts more deeply integrated symbionts. It is still biochemically independent, still carrying the full toolkit of an autonomous organism.
Ruthia magnifica is early on the spectrum. It is an intracellular symbiont, yes – committed to life inside a host – but it has not yet traveled far down what we might call the path of inevitable degradation. It is a captured bacterium whose genome still encodes the full machinery for independent life.
8.5.2 The shrinking genome: Carsonella ruddii
Now move to the other end of the spectrum.
Psyllids are small, sap-sucking insects – relatives of aphids – that feed on the phloem of plants. Plant sap is a poor diet. It is rich in sugars but deficient in essential amino acids, the building blocks that animals need to construct proteins but cannot synthesize on their own. Any insect that commits to a sap-only diet faces a nutritional crisis.
Psyllids solved this problem the way many insects have: they enlisted a bacterium. Inside specialized cells in the psyllid’s body lives Candidatus Carsonella ruddii, a gamma-proteobacterium that synthesizes the amino acids missing from the plant sap. The partnership is ancient and obligate – neither the insect nor the bacterium can survive without the other. “Successful symbiosis was the decisive factor allowing psyllids to feed only on plant sap.”14
But look at Carsonella’s genome, and you see something startling. At just 160 kilobases, it is among the smallest genomes of any known cellular organism – smaller than many viruses. It has lost genes for DNA repair, for the synthesis of its own cell wall, for most regulatory functions. It cannot make its own nucleotides. It cannot replicate without help from the host. Gene after gene has been shed, discarded as redundant once the host cell could supply the missing function.
Carsonella is so reduced that some biologists have questioned whether it should still be called a living organism. It is closer to an organelle – a piece of cellular machinery, maintained by the host, performing a specific biochemical task, unable to exist in any other context.
Genome reduction in obligate symbionts follows a ratchet-like logic. Once a function is reliably supplied by the host, the symbiont’s gene for it drifts and decays. Each loss deepens dependence, and there is no going back. Carsonella’s 160-kilobase genome illustrates a late stage.
But genome shrinkage alone does not make an organelle. That transition requires additional steps – establishment of a protein-import apparatus, coordinated gene transfer to the host nucleus, integration into host cell division – that have occurred only a few times in the history of life. Mitochondria and plastids crossed that threshold; most reduced symbionts (including Buchnera, Wigglesworthia, and others that have been shrinking for over 100 million years) have not. Genome erosion is common; organellogenesis is rare.
8.5.3 The borrowed factory: Elysia chlorotica
Between the deep-sea vent and the psyllid gut, there are stranger partnerships. Consider the sea slug Elysia chlorotica, a small, leaf-shaped mollusk that grazes on algae in the tidal marshes and shallow coastal waters of eastern North America.
When E. chlorotica feeds, it does something unusual. It punctures algal cells and sucks out the contents, digesting most of the cellular material. But it does not digest the chloroplasts – the photosynthetic organelles. Instead, it captures them intact and incorporates them into the cells lining its own digestive tract. There, surrounded by animal tissue, the stolen chloroplasts continue to function. They absorb light, split water, fix carbon. The sea slug photosynthesizes.15
The chloroplasts are not inherited. They do not reproduce inside the slug. Each generation of E. chlorotica must acquire them anew by feeding on algae. The arrangement is a temporary theft – kleptoplasty – that hints at how the permanent acquisition of photosynthesis might have begun, billions of years ago, when an ancient eukaryote engulfed a cyanobacterium and never let go.
E. chlorotica is a living thought experiment: what does the early stage of chloroplast acquisition look like? Perhaps something like this – a predator that learns to keep its prey’s machinery running, harvesting the products, and gradually becoming dependent on them.
8.5.4 The perfect commune: the three-way lichen
If Ruthia is the early stage and Carsonella is the late stage, then lichens represent something else: the fully realized partnership, stable and successful, maintained not by genomic erosion but by ecological complementarity.
A lichen is not a single organism. It is a composite: a fungus (the mycobiont) that provides the structural scaffold, intertwined with one or more photosynthetic partners. In the simplest lichens, the partner is a green alga that performs photosynthesis, converting light and CO\(_2\) into organic carbon that feeds the fungus. But in the most sophisticated lichens, there is a third partner: a cyanobacterium that fixes atmospheric nitrogen, supplying the nutrient that neither the fungus nor the alga can obtain on its own.16
Each partner retains its own genome, its own cellular identity, its own metabolic autonomy. The lichen persists not because its members have lost the ability to live alone, but because the partnership is so productive that breaking it would be a catastrophic downgrade for everyone involved.1718 The integration is shallow compared to mitochondria, but it may be the most balanced partnership on the spectrum.
8.6 Oases in the dark
The story of Ruthia is not an isolated curiosity. At hydrothermal vents and cold seeps, where reduced chemicals leak from the Earth’s interior and photosynthesis is impossible, entire ecosystems are built on symbiosis between chemoautotrophic bacteria and the animals that house them.19 The tube worm Riftia pachyptila has no mouth, no gut, and no anus; its body is packed with sulfur-oxidizing bacteria that fix carbon while the worm supplies oxygen and hydrogen sulfide via a hemoglobin that binds both simultaneously.2021 The giant clam Calyptogena hosts Ruthia in its gill cells. In every case, the animals live not by catching food from above but by partnering with bacteria that harvest the chemical energy pouring out of the Earth.
8.7 The pattern
A thread runs through these examples, though not a single trajectory. In every case, a partnership formed. In some, integration deepened and independence eroded until the boundary between host and symbiont blurred. In others – lichens, kleptoplasts – the partners remain distinct, the arrangement stable or temporary. The pattern is not a conveyor belt from symbiont to organelle. It is a landscape of possibilities, most of which stop well short of that destination.
Ruthia still has a complete genome. Carsonella has lost most of hers. Mitochondria have transferred the vast majority of their genes to the host nucleus and retained only a handful – just enough to build the core machinery of the electron transport chain, the very apparatus that made the partnership worthwhile in the first place. Chloroplasts tell the same story: once free-living cyanobacteria, now organelles with shrunken genomes, dependent on the host for most of their proteins.
The early stages of the trajectory are common: a free-living organism enters a host – by predation, by accident, by mutual convenience. If the partnership is beneficial, both partners persist. Over time, the symbiont loses genes it no longer needs, because the host supplies the missing functions. Each lost gene tightens the bond. Many symbioses stall at this stage – deeply dependent, genomically eroded, but not organelles. The final steps – evolving a protein-import system, integrating into host cell division, transferring regulatory control to the host nucleus – have occurred only a handful of times. But when they did, the line between “two organisms” and “one organism with internal compartments” vanished. That is the mechanism that built the eukaryotic cell.
8.8 The merger itself
We can now reconstruct the event – or rather, the long process – that created the eukaryotic cell.
An archaeal cell, perhaps a member of the Asgard lineage, carried genes for cytoskeletal proteins and membrane-remodeling machinery. At some point – roughly 1.6 to 2.7 billion years ago based on molecular clock estimates, with most analyses favoring about 2 billion years ago – this archaeal cell entered into an intimate association with an alpha-proteobacterium. Whether the archaeon engulfed the bacterium, the bacterium invaded, or the two grew progressively intertwined remains unclear.
The bacterium survived. Perhaps it was resistant to digestion. Perhaps the archaeal cell’s degradation machinery was incomplete. Whatever the reason, the bacterium persisted inside the host, and the two organisms began a relationship that would transform both of them beyond recognition.
The alpha-proteobacterium brought a gift: aerobic respiration. It could use oxygen – by then increasingly available in Earth’s atmosphere, thanks to billions of years of cyanobacterial photosynthesis – to completely oxidize organic molecules, extracting far more energy per glucose molecule than any anaerobic pathway could provide. The archaeal host gained access to an energy supply of unprecedented efficiency.
In return, the host provided the bacterium with a stable environment and a steady supply of organic substrates. The partnership was metabolically complementary: the host could do things the symbiont could not, and vice versa.
Over time, the two genomes began to merge. Genes moved from the symbiont to the host nucleus – a process called endosymbiotic gene transfer that continues to this day in some lineages.22 The symbiont shed genes for functions that the host could supply. The host evolved new systems for importing proteins into the symbiont, targeting gene products across the double membrane that still marks the mitochondrion as a descendant of a gram-negative bacterium.
The two partners became a single body – though not a single genome. The mitochondrion retained a small chromosome of its own, encoding core components of the electron transport chain, while the host nucleus took over the bulk of the regulatory and biosynthetic instructions.23
The result was a new kind of cell – one with a dedicated energy-producing organelle distributed throughout its cytoplasm, providing ATP wherever it was needed.
This was the birth of the eukaryotic cell.
Why did this matter for cell size? Prokaryotic cells generate ATP at their cell membrane. As a cell grows larger, its volume (which determines energy demand) increases as the cube of its radius, while its membrane surface area (which determines ATP supply) increases only as the square.24 In an influential model proposed by Lane and Martin, large prokaryotic cells face an energy bottleneck: demand outpaces supply. Mitochondria solve this by internalizing the energy-producing membranes. A eukaryotic cell can increase its volume and simply add more mitochondria, each with its own chemiosmotic membrane. The internal membrane surface area scales with volume, not with the cell’s external surface. If this model is correct, the merger did not just add a metabolic capability – it relaxed a fundamental constraint on cell size and genomic complexity.25
And then it happened again.
A eukaryotic cell – already carrying its mitochondrial passengers – engulfed a cyanobacterium. The cyanobacterium was not digested. It persisted, still photosynthesizing, still fixing carbon from CO\(_2\) using sunlight. Over time, it too lost genes, transferred others to the host nucleus, and became an organelle: the chloroplast.26
This second merger gave rise to the photosynthetic eukaryotes: the green algae that would eventually crawl onto land and become plants. Every leaf on every tree, every blade of grass, every strand of kelp carries the descendants of that engulfed cyanobacterium – a free-living organism that became a permanent component of another cell, billions of years ago.27
8.9 A merger we can still watch
Mitochondria and chloroplasts can make organellogenesis look ancient and finished. Tyler Coale and colleagues changed that view in 2024 when they showed that the nitrogen-fixing symbiont UCYN-A inside the marine alga Braarudosphaera bigelowii has crossed the threshold into something better described as an organelle: the nitroplast.28
The evidence fits an organelle in the making. The host cell encodes and imports a substantial fraction of the nitroplast’s proteins. The nitroplast divides in synchrony with the host. Its genome is drastically reduced. The partnership no longer reads as ecological convenience. It sits inside the cell’s architecture and inheritance. This appears to be a rare, modern example of primary endosymbiosis before deep time polished away the intermediate steps.29
The nitroplast matters for more than novelty. The process that produced chloroplasts was not a one-time event locked in the Precambrian. Under the right constraints, mergers can still deepen into anatomy.
8.10 The architecture of consequences
What did the merger make possible?
In the short term: larger cells with more energy, more internal organization, and the capacity to run complex biochemistry in separated compartments.
In the medium term: multicellularity. Once you have a cell with a nucleus and mitochondria, you have a cell that can specialize. You can devote some cells to digestion, others to locomotion, others to reproduction. You can build tissues, organs, bodies. The step from single-celled eukaryote to multicellular organism is not trivial, but it is a step that has been taken independently dozens of times in evolutionary history – always by eukaryotes for complex, tissue-level multicellularity – prokaryotes have evolved only rudimentary forms (filamentous cyanobacteria, myxobacterial fruiting bodies).30 The compartmentalized architecture of the eukaryotic cell is the necessary precondition for the kind of multicellularity that builds bodies.
In the long term: everything you see when you look around. Every animal, every plant, every fungus, every protist. The forests. The coral reefs. The grasslands. The humans. All built from eukaryotic cells. All carrying mitochondria. All descended from that ancient merger between an archaeon and a bacterium.
For roughly two billion years, life was prokaryotic: single-celled, small, metabolically brilliant, but architecturally constrained. The merger changed the boundary conditions. It did not violate any physical law. It did not require any new chemistry. It simply reorganized existing capabilities – archaeal information processing, bacterial energy metabolism – into a new configuration that could do things neither partner could do alone.
8.11 What competition could not build
There is a standard story about evolution that emphasizes competition: organisms fight for resources, the fittest survive, the losers go extinct. It is not wrong, but it is radically incomplete.
Competition can sharpen. It can optimize. It can hone a blade to a finer edge. But competition alone did not build the eukaryotic cell. The key innovations – aerobic respiration internalized, photosynthesis captured – came from integration: two lineages combining capabilities that neither possessed alone. The merger may well have begun as predation or parasitism rather than partnership, but the outcome was the same.
The alpha-proteobacterium did not “win” by becoming a mitochondrion. The archaeal host did not “conquer” its symbiont. Both gave up their independence. Both were transformed. And the result was not a compromise but an escalation – a cell more powerful, more versatile, and more architecturally complex than anything that had come before.
The same logic runs through every example in this chapter. The tube worm and its chemosynthetic bacteria. The psyllid and Carsonella. The sea slug and its stolen chloroplasts. The lichen’s balanced commune. In each case, integration was the creative force that competition alone could not have produced.
This is not to say that competition is unimportant. Symbiotic partnerships must still compete with other organisms and other partnerships for resources and space. Selection still acts. But the raw material that selection acts on – the new forms, the new metabolic capabilities, the new body plans – comes disproportionately from mergers.
Lynn Margulis, who spent decades championing the endosymbiotic theory against fierce resistance from the biological establishment, put it simply: “Life did not take over the globe by combat, but by networking.”31
8.12 The deep continuity
There is one more thing to notice. The merger between the archaeon and the alpha-proteobacterium was not a break in the pattern we have traced through this book. It was a continuation.
In earlier chapters, we watched prokaryotes cooperate: sharing electrons across species boundaries, forming syntrophic partnerships where one organism’s waste is another’s fuel, building biofilms where metabolic labor is divided among specialists. The logic of symbiosis – the advantage of metabolic complementarity – was already ancient when the eukaryotic merger happened.
What changed was the intimacy. In a biofilm, partners live side by side. In syntrophy, they exchange metabolites across a shared boundary. In the eukaryotic merger, one partner moved inside the other. The membrane that once separated two organisms became the double membrane of the mitochondrion – a fossil boundary, still visible under the electron microscope after two billion years.
The step from syntrophy to endosymbiosis is not a conceptual leap. It is a change in distance: from micrometers apart to nanometers apart to zero distance, to full enclosure. The driving force is the same: metabolic partnership is more efficient when transport distances are short. The closer the partners, the faster the exchange, the less energy lost to diffusion. Endosymbiosis is syntrophy taken to its logical extreme.
And the step from endosymbiosis to organelle is not a conceptual leap either. It is a change in commitment: from a partnership that could in principle be dissolved to one that cannot. Gene transfer cements the bond. Genomic erosion makes it irreversible. What was once a relationship becomes an anatomy.
8.13 You are a community
Every cell in your body contains hundreds of mitochondria. Each mitochondrion carries its own small circular genome – a remnant of the alpha-proteobacterial chromosome that has been shrinking for two billion years. That genome still encodes a few essential components of the electron transport chain, the molecular machinery that performs aerobic respiration. The rest of the mitochondrion’s proteins are encoded in your nuclear genome and imported across the double membrane after synthesis.
Your mitochondria replicate independently of your cell’s division cycle. They have their own DNA polymerase, their own ribosomes (which are bacterial-type ribosomes, not eukaryotic-type), their own translation machinery. When a cell divides, the mitochondria are parceled out to the daughter cells, not constructed from scratch. They are inherited, in an unbroken line of descent, from the mitochondria of the previous generation – all the way back, across billions of cell divisions, to the original engulfed bacterium.
If you eat a salad, the chloroplasts in the lettuce leaves tell the same story from a different chapter. They too carry their own circular DNA. They too have bacterial ribosomes. They too descend, in an unbroken line, from a cyanobacterium that was swallowed and never released.
You are a community. Not a metaphorical community – an actual one. Your cells are chimeras: archaeal information systems running on bacterial power plants. The merger is not something that happened to a distant ancestor. It is something that is still happening, right now, in every cell, in the continuous conversation between your nuclear genome and your mitochondrial genome, in the import of proteins across the mitochondrial membranes, in the division of mitochondria within your cells.
Two billion years ago, two prokaryotes merged. They abandoned cellular individuality. They combined their capabilities into a single body. And from that body came everything that followed: the algae, the plants, the fungi, the worms, the clams, the insects, the fish, the mammals, and – eventually, improbably – the chemist who now has all the test tubes she needs.
But the prokaryotic world that made the merger possible did not disappear. It continued – and continues – to run the planet’s chemistry. The syntrophic partnerships, the closed biogeochemical cycles, the layered communities we traced in earlier chapters are still at work in every sediment, every aquifer, every water column on Earth. The next chapter asks: can we write down the equation that describes what they do?
8.14 Takeaway
- Prokaryotic cells are limited by having essentially one or two internal compartments; eukaryotic cells solve this with internal membranes that create many separate reaction chambers.
- The eukaryotic cell arose from a merger: an archaeal host (likely related to the Asgard archaea) acquired an alpha-proteobacterial partner that became the mitochondrion; chloroplasts arose from a later engulfment of a cyanobacterium.
- Living symbioses – from Ruthia (early, genome intact) to Carsonella (late, genome nearly gone) – illustrate stages of symbiotic integration – partnership, gene loss, deepening dependence – though organellogenesis itself has occurred only a few times.
- Horizontal gene transfer made microbial evolution collaborative long before whole cells merged, and the 2024 nitroplast discovery shows that organellogenesis is not only an ancient event but an observable process.
- Hydrothermal vent ecosystems demonstrate that symbiosis between autotrophs and heterotrophs can sustain entire communities in the absence of sunlight.
- The creative force behind the most consequential innovations in the history of life – eukaryotic cells, photosynthetic eukaryotes, multicellularity – was integration – the merging of independently evolved capabilities – not competition alone.
Markov (2010) notes that prokaryotic cells are limited to one or two compartments (cytoplasm and periplasmic space), constraining the complexity of chemistry they can perform simultaneously. (Markov 2010)↩︎
Spang et al. (2015) reported the discovery of Lokiarchaeota from deep-sea sediments at Loki’s Castle hydrothermal vent field; the genome revealed eukaryotic signature proteins (ESPs) including actin homologs involved in membrane remodeling in eukaryotes. (Spang et al. 2015)↩︎
Spang et al. (2015) reported the discovery of Lokiarchaeota from deep-sea sediments at Loki’s Castle hydrothermal vent field; the genome revealed eukaryotic signature proteins (ESPs) including actin homologs involved in membrane remodeling in eukaryotes. (Spang et al. 2015)↩︎
The Asgard superphylum includes Lokiarchaeota, Thorarchaeota, Odinarchaeota, and Heimdallarchaeota, all named after Norse deities; see Zaremba-Niedzwiedzka et al. (2017). (Zaremba-Niedzwiedzka et al. 2017)↩︎
Eme et al. (2017) review phylogenetic evidence placing eukaryotes within the Asgard archaea, supporting a two-domain tree of life (Bacteria and Archaea, with eukaryotes as derived archaea). (Eme et al. 2017)↩︎
Masaharu Takemura, “Medusavirus ancestor in a proto-eukaryotic cell: updating the hypothesis for the viral origin of the nucleus,” Frontiers in Microbiology 11 (2020). Takemura argues that giant-virus factories provide a serious template for one route to nuclear origin. (Takemura 2020)↩︎
Karina Krarup Sand and Stanislav Jelavić, “Mineral facilitated horizontal gene transfer: A new principle for evolution of life?” Frontiers in Microbiology 9 (2018): 2217. Minerals can preserve environmental DNA and extend the time window over which horizontal gene transfer remains possible. (Sand and Jelavic 2018)↩︎
Karina Krarup Sand and Stanislav Jelavić, “Mineral facilitated horizontal gene transfer: A new principle for evolution of life?” Frontiers in Microbiology 9 (2018): 2217. Minerals can preserve environmental DNA and extend the time window over which horizontal gene transfer remains possible. (Sand and Jelavic 2018)↩︎
Molecular clock estimates for the origin of eukaryotes range from 1.6 to 2.7 Ga, with most analyses converging on ~2.0 Ga; see Parfrey et al. (2011). (Parfrey et al. 2011)↩︎
Betts et al. (2018) estimate the last eukaryotic common ancestor (LECA) at 1.84 Ga using a calibrated molecular clock. (Betts et al. 2018)↩︎
Imachi et al. (2020) cultured Candidatus Prometheoarchaeum syntrophicum, the first Asgard archaeon grown in the laboratory. Rather than a phagocyte, it was a small, slow-growing organism that extended long membrane protrusions, suggesting that the initial archaeon-bacterium association may not have required engulfment. (Imachi et al. 2020)↩︎
Newton et al. (2007) sequenced the Ruthia magnifica genome (1.16 Mb) and found it encodes a complete sulfur oxidation pathway and Calvin cycle, indicating metabolic autonomy. (Newton et al. 2007)↩︎
Newton et al. (2007) sequenced the Ruthia magnifica genome (1.16 Mb) and found it encodes a complete sulfur oxidation pathway and Calvin cycle, indicating metabolic autonomy. (Newton et al. 2007)↩︎
Nakabachi et al. (2006) reported the 160-kb genome of Carsonella ruddii, the smallest bacterial genome known at the time, lacking genes for DNA repair, cell wall synthesis, and most regulatory functions. (Nakabachi et al. 2006)↩︎
Rumpho et al. (2008) describe kleptoplasty in Elysia chlorotica, where stolen chloroplasts remain photosynthetically active for months. Early reports suggested algal genes had transferred to the slug’s nuclear genome, but subsequent genome-scale analyses found no confirmed functional transfers (Bhatt et al. 2014; Chan et al. 2018). The chloroplasts likely persist on their own long-lived proteins and mRNAs. (Rumpho et al. 2008; Bhatt and Hyman 2014; Chan et al. 2018)↩︎
Nash (2008) provides a comprehensive treatment of lichen biology, including tripartite lichens with fungal, algal, and cyanobacterial partners. (Nash III 2008)↩︎
Markov (2010) notes that prokaryotic cells are limited to one or two compartments (cytoplasm and periplasmic space), constraining the complexity of chemistry they can perform simultaneously. (Markov 2010)↩︎
Nash (2008) provides a comprehensive treatment of lichen biology, including tripartite lichens with fungal, algal, and cyanobacterial partners. (Nash III 2008)↩︎
Dubilier et al. (2008) review chemosynthetic symbioses in marine animals, emphasizing that vent and seep ecosystems are powered by bacterial primary production rather than photosynthesis. (Dubilier et al. 2008)↩︎
Riftia pachyptila lacks a mouth, gut, and anus; all nutrition is supplied by endosymbiotic sulfur-oxidizing bacteria in the trophosome; see Childress et al. (1987). (Childress and Fisher 1987)↩︎
Riftia hemoglobin has separate binding sites for O₂ and H₂S, allowing simultaneous transport of both; see Arp et al. (1987). (Arp et al. 1987)↩︎
Endosymbiotic gene transfer (EGT) moves genes from organellar genomes to the nucleus; thousands of genes have been transferred from the mitochondrial ancestor to the eukaryotic nucleus; see Timmis et al. (2004). (Timmis et al. 2004)↩︎
Lane and Martin (2010) argue that the energetic advantage of mitochondria – internalized ATP-producing membranes – gives eukaryotes on the order of 200,000-fold more energy per gene than prokaryotes, relaxing the constraint on genome size and complexity. (Lane and Martin 2010)↩︎
Lane and Martin (2010) argue that the energetic advantage of mitochondria – internalized ATP-producing membranes – gives eukaryotes on the order of 200,000-fold more energy per gene than prokaryotes, relaxing the constraint on genome size and complexity. (Lane and Martin 2010)↩︎
Lane (2005) provides an accessible account of mitochondrial bioenergetics and the surface-area-to-volume constraint on prokaryotic cell size. (Lane 2005)↩︎
Keeling (2010) reviews the origin and diversification of plastids via primary and secondary endosymbiosis; primary plastids arose once from a cyanobacterial ancestor. (Keeling 2010)↩︎
Archibald (2009) traces the evolutionary history of plastids, including multiple independent secondary endosymbiotic events in diverse eukaryotic lineages. (Archibald 2009)↩︎
Tyler H. Coale et al., “Nitrogen-fixing organelle in a marine alga,” Science 384 (2024): 217–222. The authors identify UCYN-A inside Braarudosphaera bigelowii as a bona fide organelle, the nitroplast. (Coale et al. 2024)↩︎
Ramon Massana, “The nitroplast: A nitrogen-fixing organelle,” Science 384 (2024): 160–161. A short perspective on why the nitroplast qualifies as an organelle and why it matters for understanding organellogenesis. (Massana 2024)↩︎
Grosberg and Strathmann (2007) document that complex multicellularity evolved independently at least 25 times, always in eukaryotes; prokaryotic multicellularity is limited to simple forms. (Grosberg and Strathmann 2007)↩︎
Lynn Margulis and Dorion Sagan, Microcosmos: Four Billion Years of Evolution from Our Microbial Ancestors (Summit Books, 1986). The “networking, not combat” formulation recurs throughout Margulis’s writing on symbiogenesis. (Margulis and Sagan 1986)↩︎